Chronobiologie : Principes et Méthodes - Pietro Cugini - Sémiologie Médicale et Méthodologie
Annali Instituto Superiore di Sanità, 29, 483-500, 1993
Une discipline peut être appelée
"scientifique" si elle se rapporte à un domaine concernant un
phénomène naturel qui est objectif et mesurable, probablement non causal mais
méthodique. La Chronobiologie répond à ces conditions car elle explore les
rythmes biologiques qui ont un rapport étroit avec la matière vivante. En fait,
l'objectif théorique de la recherche en Chronobiologie est la
répétitivité rythmique de phénomènes biologiques avec à
la fois l'aspect Phylogénétique (Paleochronobiologie) et Ontogénique
(Néochronobiologie).
La Chronobiologie peut être définie comme
"La discipline scientifique qui mesure et explore les mécanismes des structures temporelles
biologiques et leur relations dans les manifestations rythmiques de la matière vivante".
On doit noter que depuis que les rythmes biologiques sont génétiquement transmis,
ces phénomènes ont nécessairement un caractère hérité.
Cela implique que le temps périodique est un paramètre connivent ("counivocous") de
fonctions vivantes, qui ne peuvent pas être négligés, du fait de leur
présence dans les réflexions biomédicales traditionnelles. Les scientifiques sont
conscients du fait que les espèces vivantes vivent et agissent à la fois. Donc le
concept, du temps biologique cyclique n'est pas complètement étranger à
la théorie scientifique. La biologie traditionnelle, cependant, considère le temps
comme une quantité implicite, reléguer au rôle de facteur implicite.
L'existence et la reconnaissance de la Chronobiologie
nécessitent la renonciation au concept d'"Homéostasie", comme une
reformulation des principes biologiques établis par C. Bernard. La Chronobiologie soutient que
la Physiologie des fonctions vitales ne répond pas aux lois de l'état stabilisé,
invariant, où l'inconditionnel retour à l'équilibre initial est du à
une perturbation. Inversement, la répétition du rythme de phénomènes
biologiques démontre l'idée que la matière vivante est sujette à une
continuelle variation d'état. En conséquence, les rythmes biologiques représentent
un processus syntropique-entropique avec accumulation et dissipation d'énergie vitale. Dans les
mécanismes biopériodiques la matière vivante trouve le temps nécessaire
pour la consommation et la reconstruction de matériaux organiques.
CLASSIFICATION DES RYTHMES BIOLOGIQUES
Les rythmes biologiques peuvent être classés selon de nombreux critères.
1. Classification physique
Cette classification est basée sur la longueur de la période
d'oscillation (voir table 1)
TABLE 1 Classification temporelle des rythmes biologiques
--------------------------------------------------------
Domaine :.. Période (Tau)
--------------------------------------------------------
Ultradien : <20 h
Circadien : 24 +/- 4 h
Dien : 24 +/- 2 h
Infradien : >28 h
Circaseptien: 7 +/- 3 j
Circadiseptien : 14 +/- 3 j
Circavigintien : 21 +/- 3 j
Circatrigintien : 30 +/- 5 j
Circannuel : 1 a +/- 2 m
-------------------------------------------------------
h = heures j = jours m = mois a = année
Les rythmes qui ont une période d'oscillation de 24 +/- heures sont définis comme
"Circadien" (de Circa dies, c'est à dire, à peu prés un jour). Les
événements cycliques avec une période de moins de 20 heures ou plus de 28 heures
sont dit respectivement "Ultradiens" et "Infradiens".
2. Classification fonctionnelle
En plus de classification physique il existe une subdivision basée sur des concepts fonctionnels
qui reconnaissent quatre variétés de rythmes biologiques, c'est à dire, Alpha,
Bêta, Gamma et Delta.
Les rythmes Alpha coïncident avec les oscillations
spontanées des fonctions biologiques. Le rythmes Alpha sont sous divisés en Alpha(s) et
Alpha(f) selon qu'ils soient produits dans des conditions de "synchronisation" ou de "mouvement libre"
("free-running")(voir plus loin). Les rythmes Bêta correspondent à la
périodicité de la réponse des fonctions biologiques en cas de stimulations ou
d'inhibition pour différents temps. Les Bêta rythmes existe aussi en
variété Bêta(s) et Bêta(f) en relation de la présence soit de
conditions de synchronisation ou de "mouvement libre" (free running). Ces deux variétés
sont sous divisés en Bêta(s1) ou Bêta (f1) si la perturbation est physiologique, et
Alpha (s2) ou Bêta (f2) si la perturbation n'est pas due à un phénomène
physiologique. Les rythme Gamma concernent les oscillations périodiques de fonctions biologiques
qui sont modulées, perturbées ou influencées par des facteurs
déterministes soit physiologiques, c'est à dire, Gamma (s1) ou Gamma (f1), ou non
physiologiques, c'est à dire, Gamma (s2) ou Gamma (f2). Ici à nouveau,
la différenciation entre les variétés Gamma(s) et Gamma(f) dépendent de la
présence soit de conditions de synchronisation ou de mouvement libre (free-running). Enfin,
les rythmes Delta, qui sont sous divisés en variétés (s) et (f), correspondent
à la modification dans l'oscillation périodique d'une fonction biologique
donnée secondaire de la manipulation d'un rythme Alpha, Bêta ou Gamma.
3. Classification mathématique
L'examen de phénomène rythmique dans la matière
organique révèle qu'il existe des événements qui se répètent
après un certain laps de temps à des occurrences isolées. Il y a des
"rythmes qualitatifs, ponctuels, discrets ou épisodiques" qui expriment par une condition
binaire, c'est à dire, présent ou absent, selon un événement ou un
non-événement. Par exemple le cycle menstruel.
Les rythmes qualitatifs sont descriptibles mathématiquement en terme de quantités finies
(0 ou 1) et comptés comme des fréquences numériques. Ainsi, les rythmes
qualitatifs peuvent s'appeler aussi "rythmes fréquentiels".
Dans les organismes vivants on peut remarquer que plusieurs
phénomènes se répètent comme des événements qui varient dans
un "continuum". C'est à dire, que le phénomène est toujours
présent et mesurable, bien que changeant en fonction du temps. Sa grandeur garde le
même niveau suivant une période de temps donné. Ainsi, la période de ce
phénomène est donnée par sa durée dans laquelle la courbe atteint le niveau
identique après une oscillation complète. Ces événements périodiques
ont ainsi une expression quantitative si leur variabilité peut être identifiée
comme "des rythmes analogiques ou continus ou quantitatifs". Ces rythmes s'expriment
mathématiquement par des valeurs d'un ordre potentiellement infini.
D'un point de vue de classification, il existe un troisième
type de rythme biologique consistant en des pics isolés sur la courbe d'oscillation
quantitative. Si ces sauts montrent une cadence dans le temps, ils peuvent être définis
comme "rythmes épisodiques".
4. Classification descriptive
Cette classification est principalement utilisée pour la
description de rythmes épisodiques ou quand il est nécessaire de décrire un
événement périodique en relation avec ces pics. Les rythmes inclus dans ce type
sont diurnes, nocturnes, "serotine", vespérales, matinaux, journaliers, hebdomadaires,
mensuels, saisonniers, annuels, etc.
On peut noter, cependant, que ces conditions définissent la
périodicité de manière descriptive et ne s'ajoutent pas à une quelconque
conclusion sur la durée effective de la période du phénomène
récurrent. Ainsi, un rythme diurne n'est pas implicitement Circadien ; il peut être
Ultradien.
5. Classification évolutive
Les rythmes biologiques, comme tous les phénomènes
biologiques, passent par un processus évolutif qui a tendance à modifier les
propriétés périodiques en fonction de l'âge chronologique. Comme dans la
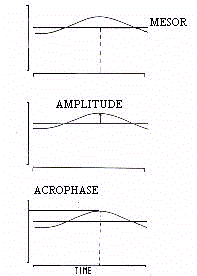
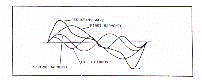
figure 1, chaque fonction périodique est définie par son niveau moyen, la mesure de son
oscillation, le temps de sa crête oscillatoire, ces paramètres sont appelés
respectivement, Mésor (M), Amplitude (A) et Acrophase (ø ou Phi).
En utilisant la méthode du Clinospector (voir plus loin), les signaux positifs ou
négatifs sont identifiés par leur Mésor et leur amplitude et correspondent
à un effet de l'âge (Fig. 2).
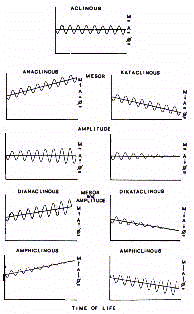
Ainsi il y a des rythmes Déclinants ("Dianaclinous" ou
"Dikaclinous") si leurs propriétés ont une tendance positive ou négative pendant
le cours de leur vie, si l'évolution à travers le temps chronologique concerne un seul
de ces paramètres dans un sens positif ou négatif. Il y a aussi la possibilité
d'un tendance opposée aux deux propriétés rythmiques définies comme
rythme Amphiclinique ("Amphiclinous"). Enfin, il peut s'agir d'un rythme Aclinique ("Aclinous") qui est
un rythme qui reste stable même avec l'augmentation de l'âge.
6. Classification en durée
En considérant les rythmes biologiques, on peut constater
que certain sont "permanents ou avec une longue durée" et d'autres "transitoires
ou temporaires". Le cycle ovarien est un rythme typiquement "provisoire" par ce qu'il disparaît
avec la ménopause. Le rythme de la température du corps est par contre un rythme
permanent que l'on peut toujours être observé dans les 24 heures après la mort.
7. Classification physiologique
Si on considère les rythmes biologiques, on ne peut pas
négliger le rôle important qu'ils jouent dans les fonctions vitales. Dans ce sens il doit venir à l'esprit
qu'il y a des rythmes "essentiels" ou "vitals", et des rythmes "non-essentiels". Des rythmes essentiels sont par exemple
l'activité pulsative du coeur, de la respiration, et de l'activité électrique
cérébrale. La suppression de l'un correspond à la "mort physique".
L'absence du rythme cérébral, comme on peut le voir sur un
électroencéphalogramme défini la "mort clinique".
Nous pouvons en déduire que, d'après ce que nous
avons dit, les rythmes essentiels représentent la vie biologique. La mort correspond à
l'annulation de l'existence de ces rythmes fondamentaux. De ce point de vue on voit que la vie et la
rythmicité biologique sont des expressions qui se ressemblent de manière connivente
("counivocous").
Les rythmes non essentiels sont les rythmes dont la suppression
ou la désynchronisation n'a pas de répercussion sur les fonctions vitales.
8. Classification biologique
Vu sous l'espect de leur signification, les rythmes biologiques
peuvent être trouvés dans deux aspects importants de la vie, c'est à dire, dans
les fonctions conservatrices et reproductrices.
Les rythmes du domaine conservateur sont, par suite, mentaux et
physiques. Ces deux catégories sont intellectuelles, affectives, endocrines cardio-vasculaires,
métaboliques, respiratoires, digestives, etc. Les rythmes reproducteurs sont, d'autre part,
relatifs à la sexualité et la fertilité.
9. Classification selon la résistance
En biologie, la manifestation rythmique de la vie a
différentes valeures en fonction de sa robustesse vitale.
Il existe, en réalité, des rythmes "résistants"
ou "stables" en plus des événements biopériodiqes qui sont "faibles" ou "labiles".
La résistance (dans le sens robustesse) d'un rythme dépend de son rôle et de quel
système il appartient. A la base, la résistance est inversement proportionnelle à
la susceptibilité d'être désynchronisé suivant des perturbations
importantes (graves).
La labilité d'un rythme est en majorité
dépendant de son passage spontané ou forcé à un ordre différent de
périodicité (multiplication ou démultiplication de fréquence). Le rythme
du coeur est dit labile par ce qu'il peut facilement varier en fréquence au cours d'une
durée de 24 heures. Rarement la labilité d'un rythme est due à son abolition.
L'abolition d'un rythme biologique est un événement extrêmement non naturel
qui est très improbable. Ainsi, la disparition d'un rythme donné peut être
prudemment évaluée et ne peut pas être survenu sans avoir vérifié
que le rythme a juste et simplement changer de période.
10. Classification ontogénique
Les rythmes biologiques sont une partie du patrimoine
génétique de la matière vivante. Les oscillateurs sont localisés dans
chaques cellules, à n'importe quel niveau d'organisation. Les rythmes commencent à
agir à la naissance de la cellule. Chez les métazoaires les rythmes de la cellule
prennent part à rythmicité très complexe et générale dont
l'expression nécessite coordination et maturation. Quelques rythmes de hautes activités
organisées, ainsi, prennent un certain temps pour leur Ontogenèse postnatale.
Les rythmes dans des phases de formation sont dits rythmes
"immatures", tandis que ceux qui opèrent à la naissance sont dits rythmes "matures".
11. Classification structurale
Les rythmes biologiques sont des événements naturels qui reviennent
spontanément, leurs composants périodiques sont endogènes. Leur contribution
endogène à la périodicité peut se manifester librement
("free-running rhythms") ou peut être conditionnée par des facteurs d'environnement
qui sont cycliques comme des synchroniseurs (rythmes synchronisés et masqués). Les
rythmes à mouvements libres ("free-running rhythms"), donc, peuvent être
transformés en rythmes synchronisés, et le point final de cet effet est un " effet
masquant " exercé par un composant exogène sur une bio-périodicité
endogène. Dans la nature, la manifestation évidente pour la plupart des rythmes
biologiques est la combinaison de composant endogène plus un entraînement
exogène. Dans ce cas, l'effet masquant résulte en un rythme synchronisé,
et les facteurs externes du masquage peuvent être définis comme des
"agents entraînants" ou "zeitgebers" ou "synchroniseurs".
Important, la manipulation d'un synchroniseur environnemental peut
provoquer le dérangement de la périodicité endogène qui consiste en un
phénomène de "désynchronisation externe". L'effet non chronique
doit s'appliquer quand on fait référence à une périodicité
manquante du rythme biologique. Cela veut dire que la suppression d'une périodicité
donnée peut être attribuée à un mécanisme exogène non
principalement dépendant d'un défaut intrinsèque d'une rythmicité
interne.
De façon plus intéressante, l'effet masquant peut être non seulement
exogène mais aussi endogène. L'effet masquant endogène peut expliquer les
conditions complexes d'une perte de rythme non explicable en terme de cause à effet.
Par exemple, la perte d'un rythme sommeil-éveil produit un effet masquant endogène
sur de nombreux autres rythmes provoquants l'annulation de leur périodicité
(voir plus loin).
12. Classification selon la cohérence ("consistency classification")
Il y a des rythmes qui concerne une entité concrète,
c'est à dire, les "rythmes réels". D'autres rythmes sont au lieu de cela l'expression
unique d'un paramètre calculable, c'est à dire des "rythmes virtuels". Le profil
nycthéméral du rythme du Cortisol, quand il est étudié dans le sang,
coïncide avec les variations journalières de sa concentration dans le plasma ou le
sérum. Par contre, le rythme circadien du pH est causé par l'effet de nombreux facteurs
chacun caractérisé par son propre rythme.
13. Classification constitutive
Il y a des rythmes biologiques qui se référent
à une seule variable (quelques chose qui est défini par ses caractéristiques),
c'est à dire des "rythmes composites". Par exemple le rythme circadien de la prolactine,
d'une part, et le rythme circadien de l'humeur ("mood") d'autre part.
Si un rythme est défini comme complexe, sa suppression éventuelle
peut être dépendante d'une désynchronisation interne parmi les facteurs cycliques
ou des mécanismes. Quelques fois, la périodicité est seulement due aux changements
de phase résultant d'une oscillation antiphasique des cycles qui contribuent au rythme
complexe.
14. Classification hiérarchique
Dans la magnifique organisation des phénomènes
bio-périodiques on a trouvé que quelques rythmes jouent un rôle important dans le
conditionnement d'autres cycles biologiques. Ces rythmes sont appelés "guide ou rythme primaire
ou indépendant" pendant que les rythmes dirigés ("driven rhythms") sont appelés
"rythmes guidés ou secondaires ou dépendants". Les rythmes guidés ont une
importance stratégique dans le sens que leur présence est essentielle pour les
périodicités dépendantes. Ce qui manque à un rythme guidé
c'est qu'il produit des effets désynchronisés du à la suppression de l'effet
rythmique. La suppression de la relation entre les rythmes primaires et secondaires cause un
phénomène appelé "désynchronisation interne". Les rythmes guidés
disparaissent par un effet induit. De même qu'une répercussion dramatique est
appelée "effet masquant endogène". Le masquage endogène nous aide à
comprendre et à interpréter la Chronopathologie de certains rythmes biologiques.
GENESE DES RYTHMES BIOLOGIQUES
La capacité de subir des oscillations rythmiques est une
caractéristique intrinsèque de la matière vivante. Un fondement important de la
Chronobiologie est que "de nombreux rythmes persistent même dans le cas d'une isolation
complète pour la plupart des cycles de l'environnement". Cette affirmation clarifie que les
rythmes naturels peuvent être considérés comme en dehors des cycles
géophysiques. Cela veut dire que la matière vivante a son propre temps, c'est à
dire, le "temps biologique".
En considérant le temps comme la quatrième
dimension en biologie, on peut conceptuellement et de manière syllogistique dire qu'un
Chronome existe dans le Génome. En outre les signaux physiques (physèmes) et chimiques,
on peut affirmer que les gènes fournissent aussi de l'information sous forme de
"chronèmes", c'est à dire, des signaux de type périodique. Ainsi le processus
de clonation (duplication) est temporisé par des périodes déterminées,
et résulte d'une combinaison de messages quantals (quantal) et temporelles qui pousse
quantitativement les fonctions biologiques à des changements selon un spectre programmé
de périodicités. Théoriquement, on peut supposer que la genèse
périodique des signaux temporels trouve son origine dans les spirales hélicoïdales
du DNA là où le Chronôme doit résider. La double hélice du DNA peut
agir comme un métronome générant une vibration dont la longueur est la
période de clonation (duplication).
HORLOGES BIOLOGIQUES ET CONTROLE DU PHENOMENE BIOPERIODIQUE
Les périodicités biologiques sont conduites par un programme biologique selon une
durée temporelle (biotemporality) qui cause une récursion dans le spectre de
fréquence allant de la millisecondes à des années.
L'effet temporel de la programmation génétique,
le chronome, est le composant endogène pour lequel les rythmes biologiques se produisent
comme des événements à "mouvements-libres" (free-running). Les rythmes
à mouvements libres reflètent le "temps du corps" qui est indépendant du temps
de l'environnement mesuré par une horloge, le "temps physique". Les rythmes à
mouvements-libres reflètent les mécanismes endogènes d'une temporisation
cyclique dont la manifestation est morphologiquement comme une horloge interne, une "horloge
biologique".
En considérant les animaux dans leur environnement, on peut noter que les
rythmes endogènes sont souvent à mouvements libres. Le "temps du corps" est
masqué, et les rythmes biologiques spontanés sont obligés par les cycles
exogènes d'ajuster leur période pour être en accord. Cela veut dire que le temps
biologique a de façon innée la capacité de s'uniformiser avec le temps physique.
Donc, les événements qui perturbent l'environnement peuvent moduler la cadence
périodique issue de leur hérédité détermine des rythmes
endogènes. Les fortes interférences sont celles qui sont fournies par des
événements systématiques qui possèdent un caractère cyclique dans
leur manifestation. L'alternance lumière-obscurité, le temps de l'alimentation,
les routines sociales, en incluant les décalages du travail, etc. (voir plus loin) sont des
agents entraînants déterminants.
Dans le cas d'untraînement de rythmes endogènes de
nombreuses entités structurales interviennent avec un rôle de médiation
(Table 2).
TABLE 2 - Structures nerveuse centrales impliquées dans
la chronorégulation de fonctions biologiques.
-------------------------------------------------------------
Noyau suprachiasmatique
Bulbe olfactif
Fornix
Septum
Structure limbique
Hippocampe
Zone préoptique
Connection rétino-hypothalamique
"Midbrain raphe nuclei"
Hypothalamus ventromédian
Hypothalamus dorsomédian
Locus coeruleus
Pédoncule cérébral
Système nerveux autonome
Ganglion supérieur cervical
Glande pinéale
-------------------------------------------------------------
Les déterminants les plus importants des synchroniseurs
biologiques sont les oscillateurs endogènes, ce sont des structures de l'organisme qui
fonctionnent comme des "pacemakers" rythmiques. D'autres dispositifs de synchronisations sont
les "space-resetters;", éléments de l'organisme qui régulent la structure
temporelle d'un ou plusieurs rythmes en réponse à un ou plusieurs synchroniseurs
environnementaux. Les relations entre les pacemakers et les pace-resetters sont
déterminées par des structures de connections spéciales appelées
"tranceducers" qui traduisent le stimulus exogène aux horloges internes. Les transducteurs
peuvent avoir un effet soit négatif, soit positif sur les oscillateurs. Les séries
peuvent être intégrées par les "modificateurs" ou les "contrôleurs
logiques" qui agissent comme, respectivement, en modifiant et contrôlant les stimulus
exogènes et endogènes.
Concernant les horloges biologiques il existe une éternelle
polémique entre les positivistes et les négativistes. L'opinion dominante est celle
des positivistes qui disent que les horloges biologiques sont des entités identifiables qui
se localisent dans les tissus d'organes. Ils croient dans l'existence d'horloges biologiques et
soutiennent que ces structures de maintenance de synchronisation jouent un rôle dans la
coordination d'une grande quantité de rythmes biologiques périphériques.
Une telle capacité coordinatrice suppose un commandement avec lequel les horloges
biologiques conduisent la période des rythmes provenants de chaque cellule de
l'organisme. Cela implique que les horloges biologiques sont susceptibles d'interagir avec toutes les cellules
en connaissant les messages nerveux, physiques et chimiques. Pour cette raison ils sont
localisés à l'intérieur des structures non mitotiques du système
nerveux, encéphalique et spinal.
L'organisation structurale des horloges biologiques est
difficile à déchiffrer. Une tentative peut être faite en présentant les
principaux modèles. Le modèle (I) le plus simple, est formé d'un oscillateur
qui synchronise un second oscillateur, et ainsi de suite. Ce modèle fondamental propose un
contrôle cybernétique linéaire. Le modèle (II) décrit un premier
oscillateur suivi par successivement une série d'oscillateurs. Le modèle (III) propose
une interaction entre plusieurs oscillateurs d'importance hiérarchique égale
ordonnés dans un réseau cybernétique. Ce mécanisme interactif de
chronorégulation est appelé "feed-sideward".
La recherche en Chronoanatomie a permis de mettre en
évidence des structures responsables de la programmation rythmique et l'intégration
Chronobiologique de l'organisme avec l'environnement. L'information des structures neuroanatomiques
sont impliqués dans la régulation centrale des rythmes biologiques provenant
essentiellement de l'étude animale. La table 3 liste les structures qui sont effectivement
reconnue comme jouant un rôle rythmogénique d'oscillateur.
Table 3 : Structures centrales impliquées dans la coordination des fonctions oscillatoires
biologiques.
-------------------------------------------------------------
Pacemakers
Paceresetters
Modifiers
Transducers
Logic-controllers (contrôleurs logiques)
Synchronizers (synchroniseurs)
-------------------------------------------------------------
DESYNCHRONISATION, RESYNCHRONISATION, CHRONISATION
Les travaux en Chronobiologie ont mis en évidence que
les facteurs environnementaux agissent hiérarchiquement comme des synchroniseurs de rythmes
biologiques. Le synchroniseur le plus puissant est l'alternance lumière-obscurité.
Isolé du temps géophysique, l'être humain à tendance de retarder le temps
de repos. Ce phénomène arrive même dans les conditions de lumière
constante ou d'obscurité. Un changement rapide dans les zones temporelles (passant de trois
zones temporelles ou plus), arrive dans les vols transméridiens, qui donne lieu à
une perturbation psychophysique connue sous le nom de "jet lag syndrome" due au
désynchronise entre le temps biologique et le temps physique. La resynchronisation suit le
désynchronise géographique qui suit arrive avec un décalage de phase d'à
peu près 90 minutes tous les 24 heures. C'est, pourtant, nécessaire de garder en
tête que la direction de la transition de zone temporelle est cruciale. Dans les vols Est-Ouest,
les voyageurs doivent récupérer un temps égal à la différence
temporelle entre les deux zones de temps. Dans les vols Est-Ouest, les sujets doivent
récupérer 24 heures moins la différence des zones de temps,
c'est à dire, le temps physique passé dans la zone qui n'a pas été
vécu " lived " par les voyageurs. Cela implique que la resynchronisation prend
beaucoup plus de temps. Cependant la resynchronisation peut être accélérée
ou retardée par de nombreux facteurs (table 4)
Table 4 : Facteurs affectant la vitesse de resynchronisation.
Accélération Retard
---------------------------------------------------------------------------------
Extraversion Introversion
Seralité Manilité
Avance de phase Retard de phase
Décalage en retard Décalage en avance
Rythme labile Rythme stable
Pression temporelle forte Faible pression temporelle
Tache de haute performance Tache de basse performance
Jeunesse Vieillissement
Névrose basse Haute névrose
Faible pulsation/ Pulsation rapide/
coefficient de respiration faible coefficient de respiration élevé
--------------------------------------------------------------------------------------
Un repas est aussi un synchroniseur important. Des sujets
alimentés avec un repas complet, une fois par jour montre un décalage de phase pour
de nombreux rythmes biologiques suivant l'heure ou le repas sont pris. Des procédures sociales
(socio-temporalisme) sont important aussi, spécialement un décalage dans le travail.
Un décalage aléatoire peut produire des effets de désynchronisme pour de
nombreuses fonctions périodiques, spécialement pour les performances mentales ou
physiques. D'autres agents d'environnement sont responsables de désynchronisation comme le
stress, la vitesse, la fatigue, etc., s'ils sont anormalement prolongés dans le temps ou
répétés d'une manière cyclique.
Diverses drogues peuvent induire une désynchronisation.
La liste de ces drogues pourrait composer un nouveau chapitre de Pharmacologie et être
utilisé en surveillance pharmacologique (pharmaco-vigilance).
Certaines drogues peuvent être utilisées pour
resynchroniser les rythmes biologiques lorsque ceux-ci sont perturbés par des
interférences exogènes. Ces drogues sont appelées " agents chronisant ou
chroniseurs ". Une liste de molécules pharmacologiques favorisant la chronisation sont
donnés dans la table 5.
Table 5 : Agents pharmacologiques utilisés comme drogue Chronobiotique.
ACTH
Barbituriques
Antidepresseurs tricycliques
Lithium
Nomifensine
Dérivés de la Xantine
Levodopa
Dépleteurs de la 5-hydroxytryptamine
Indométhazine
Mélatonine
-----------------------------------------------------------
ONTORYTHMOGENESE ET GERORYTHMOCLINIE
Etant l'expression du dynamisme vital, les rythmes biologiques
commencent au temps de la fécondation, appelé " Point zéro
Ontogénétique ". Certains rythmes nécessitent une coordination centrale
contrôlée par des horloges biologiques dont le développement est relatif
à la maturation du système nerveux central.
Le besoin de maturation montre que l'antogenèse des rythmes
biologiques résulte du recrutement, de l'organisation et de la synchronisation d'un grand
nombre de cycles générés par des cellules au niveau
périphérique.
Il est important, toutefois, de remarquer que certains rythmes
requièrent un haut degré d'untégration, d'organisation et peuvent être
retrouvés chez le fétus. La synchronisation fétale renforce l?dée que les influences
maternelles de cycle naturel sont actives dans l'organisation temporelle de fonctions
biopériodiques in utero.
Après la période de maturation, quelle est la
destinée des rythmes biologiques considérés comme fonction de l'âge
chronologique ? En d'autres termes, quel est l'effet du vieillissement sur les rythmes
biologiques ?
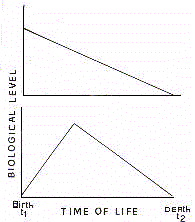
En biologie traditionnelle, le processus de vieillissement
consiste en un phénomène de perte linéaire des fonctions biologiques
(Fig 3)
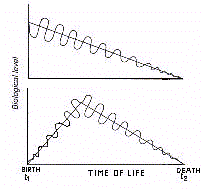
La Chronobiologie considére le vieillissement, dans le
contexte de cyclicité qui caractérise les fonctions biologiques. Des travaux ont
montrés que la senescence est associée avec une déclinaison qui affecte le niveau
moyen (déclinaison gérontologique de l'activité tonique), et l'étendue
(déclinaison gérontologique de l'activité phasique) de l'oscillation, en plus
d'un décalage dans la phase (rephasement gérontologique) et/ou un changement dans la
période (démultiplication/multiplication gérontologique) des rythmes
biologiques. Ainsi on peut dire que le vieillissement est un processus associé
à une direction (ascendante) avec une hauteur négative et une forme oscillatoire de
fonctions biopériodiques. Cela implique que les effets de sénescence consiste en un
modèle de régression périodique linéaire, dans laquelle on peut voir
une chute des oscillations dans son niveau moyen et d'amplitude, c'est à dire,
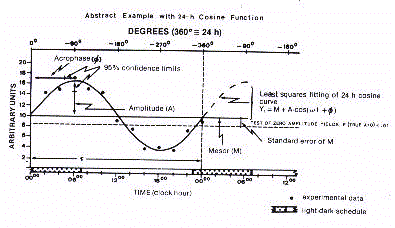
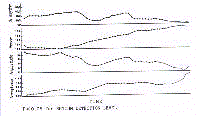
un phénoméne de gérorythmoclinie (Fig. 4)
A présent, l'analyse de régression périodique
linéaire de la tendance gérontologique des rythmes biologiques peut s'effectuer
par la méthode du Clinospector. Cette procédure, développée par l'auteur,
combine l'estimation des changements oscillatoires, période par période, comme une
fonction de l'âge avancé linéaire. Le modèle du devenir
gérontologique des rythmes biologiques est représenté par une courbe,
c'est à dire, le gérorythmoclinogramme (Fig. 2)
Ainsi, la clinorythmométrique étudie la
sénescence des rythmes biologiques qui peuvent montrer une déclinaison (rythmes
katacliniques ou katarythmes), d'autres peuvent montrer une amplification (rythmes anacliniques
ou anarythmes), dans laquelle la moyenne de l'amplitude de l'oscillation, et finalement, d'autres
peut apparaître non pas du tout comme un changement (rythmes ascendants ou isorythmes), avec un
âge avancé. Le gérorythmoclinogramme de cette tendance est
représenté par la figure 2.
La tendance kataclinique est une expression typique des rythmes
"causatifs" de sénescence. La tendance anaclinique, par contraste, représente le
modèle périodique linéaire des rythmes "adaptatifs" de la vieillesse.
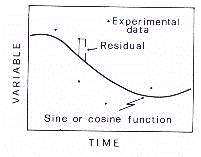
DEFINITION D'UN RYTHME BIOLOGIQUE
Dans les paragraphes précédents,
les phénomènes périodiques ont été traités comme des
signaux temporels dont la variation peut être exprimée par des valeurs discrètes
coïncidants avec le temps de l'échantillon.
On doit insister sur le fait qu'une série discrète
temporelle n'est pas une fidèle représentation du signal dans le fait que
l'approximation dépend du temps et de la densité des données collectées.
(Fig. 5).
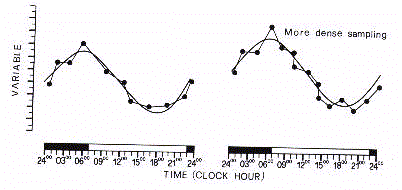
Dans la discrétisation des phénomènes
biopériodiques il y a un risque d'orreurs systématiques, dans toute série
temporelle constituée par un composant intrinsèque qui est superposé par des
éléments occasionnels ou aléatoires. Les séries discrètes peuvent,
donc, être contaminées par des valeurs aberrantes, appelées " bruit
biologique ", n'appartenant pas au composant systématique de la variation
périodique (Fig. 6).
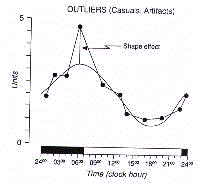
Afin d'unlever le bruit biologique du aux valeurs anormales,
appelées "outliers", le signal périodique doit être approximé
géométriquement par une courbe ajustée à la série de
données (Fig 7.)
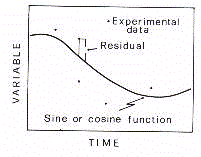
Comme le signal est périodique, la courbe analytique doit
être représentative de l'oscillation harmonique. Donc, en supposant le rythme biologique
est mieux représenté par les ondes oscillatoires qui calculent le signal discret dans
ces fluctuations périodiques. On peut suggérer que le rythme biologique est par
définition "le composant périodique d'une série temporelle de données
biologiques dont le profile d'onde a été validé analytiquement".
En d'autres termes, un rythme biologique est "un biosignal dont la période d'oscillation a
été validée par des modèles analytiques de régression
périodique utilisant un ou plusieurs composants harmoniques". On peut dire qu'une variation
discréte des données biotemporelles n'est pas implicitement un modèle
représentant un rythme biologique. Manifestement, la résolution analytique d'un
biosignal discret n?mplique pas que le rythme biologique est représenté par une forme
sinusoïdale parfaite. La "sinusoïdalisation" est juste un modèle pour enlever les
composants aléatoires qui sont responsables des bruits biologiques, et ainsi de l'erreur
biométrique.
De plus, il y a d'autres raisons biostatistiques qui
reconnaissent la représentation harmonique comme un signal idéal de cycles biologiques.
Une variation arrivant après une échelle de 24 heures n'est pas une démonstration
implicite que le modèle varie selon une période de 24 heures, il y a un besoin de valider
statistiquement l'oscillation dans sa périodicité. La niveau de détection du
rythme peut être statistiquement vérifiée en supposant une probabilité
(P) "alpha" de moins de 5% pour l'hypothèse nulle de non oscillation (amplitude zéro).
En utilisant le modèle harmonique, la rythmicité biologique peut être, ainsi,
validée statistiquement dans sa significativité contre la probabilité de P
< 0.05 que l'effet d'oscillation est due aux effets du hasard.
Les méthodes en régression périodique
utilisées dans la détection analytique et le test de sinusoïdalité de
séries temporelles discrétes de données biologiques sont abordées dans le
chapitre suivant.
METHODOLOGIE EN CHRONOBIOLOGIE OU CHRONOBIOMETRIE
Dans les séries temporelles traitées en
Biométrique de données biologiques, le temps (T) peut être considéré
comme un paramètre systématique qui est indépendant de la variable biologique que
l'on étudie. Le temps explique comment les biovariables changent, mais ce n'est pas la cause
déterminante de ce changement, qui, par suite, dépend des propriétés
dynamiques de l'entité biologique (au sens vivante)
Statistiquement parlant, un phénomène Y
et un temps T sont liés, mais le temps n'est pas une variable aléatoire, sa variation
est numériquement prédictible.
En général, la valeur de Y (Yt) pour un
temps donné (t) est liée à la valeur précédente. Pour cette raison,
une série temporelle peut être qualifiée d'autorégressive et historique. Si
la relation entre Y et T préserve celui-ci inchangé, nous avons là
un phénomène temporel linéaire. Si la relation change spontanément nous
avons affaire à un événement temporel non linéaire. Si la relation reprend
la même valeur à un temps t nous avons affaire à un phénomène
cyclique.
En termes descriptifs, une donnée temporelle
d'un phénomène cyclique, basée sur une unité donnée de temps peut se
diviser en divers composant : 1. Un composant harmonique fondamental qui exprime la structure
périodique d'un phénomène rythmique ; 2. Un ou plusieurs composants
sous-harmoniques dont la période est un sous-multiple de la composante principale ; 3. Un
composant aléatoire qui correspond au bruit éliminé par la courbe oscillante.
Ainsi, le bruit biologique est caractéristique
d'une série discrète, et il influence les estimations biométriques des
statistiques numériques. Toutefois, le bruit biologique n'est pas responsable dans les
séries continues, et n'a pas qu'une petite influence dans les statistiques analytiques. Dans les
séries de données temporelles analysées biométriquement, la chronobiologie
applique deux méthodes, c'est à dire, 1. Numérique ou non déductive ou
Chronobiométrie macroscopique ; 2. Analytique ou déductive ou microscopique
Chronobiométrie.
I. CHRONOBIOMETRIE NON-DEDUCTIVE (ANALYSE MACROSCOPIQUE)
La Chronobiométrie non déductive est basée
principalement sur des mesures d'emplacement principal et de la dispersion des données
expérimentales à chaque point temporel (Fig. 8)

La représentation des valeurs moyennes
résultantes de cette analyse est appelée "Chronogramme".
La dispersion de chaque point autour de la moyenne peut
être mesurée par l'écart-type (SD), écart-type de la moyenne (SE) ou (SEM)
ou limites de confiance à 95% (95%CL).
L'effet véritable du temps sur une série
temporelle de données peut être constaté par une "One-way variance analysis"
(One-way ANOVA). La constance (homogeneity) de la variance en chaque point ("homoschedasticity") peut
être vérifiée par la connaissance du calcul du test de Bartlett ou du test de
Duncan.
Les valeurs estimées dans le Chronogramme
sont : 1. Le niveau moyen global (overall mean level); 2. Le niveau du maximum (peak level);
3. Le niveau du maximum du temps (peak time) (zenith) ; 4. Le niveau de la dépression
(trough level) ; 5. Le temps de la dépression au minimum (nadir) ; 6.
La différence entre les maximums et les minimums (difference between peak and trough
levels) ; 7. Les coefficients de variation des niveaux moyens du temps modéré
(coefficient of variation for time-qualified mean levels)
II. CHRONOBIOMETRIE DEDUCTIVE (ANALYSE MICROSCOPIQUE)
L'approche analytique des séries temporelle comprend
diverses méthodes.
A. Chronobiométrie déductive avec période inconnue.
1. Analyse de la meilleure période calculée (périodogramme)
Dans ce type d'analyse la meilleure forme d'onde calculée
est trouvée en utilisant la fonction
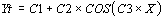
Le coefficient le plus important dans cette formule est C3 et
qui correspond à la période qui approxime de façon optimale les séries
temporelles de données.
2. Analyse Spectrale
L'analyse de la résolution spectrale (Fourier analysis)
est une méthode des plus importante dans le traitement numérique des signaux quand ils
sont discrets (signaux numériques) ou continues (signaux analogiques).
Le signal discret pour la résolution dans les
séries harmoniques (Transformation de Fourier Discrète) peut être
représenté par X termes équiréparties dans le temps.
La formule est :
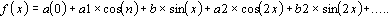
La relation entre le numéro de terme (n) et le temps (t) est :
t = nI
Ou I est l'intervalle d'aeacute;chantillonnage. L'intervalle entre
les valeurs étant donné, t correspond au temps d'un cycle ou période (TAU) ou
durée du signal.
L'analyse spectrale consiste à la transformation du
signal séquentiel, c'est à dire, un entité temporelle, en spectre de
fréquence, c'est à dire, une entité de fréquence. Le spectre des
fréquences calculées est la microstructure du signal décomposé (Fig. 9).
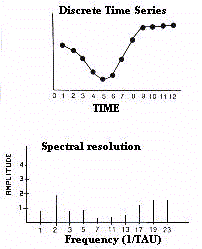
En posant la période égale à TAU,
la formule pour l'analyse spectrale peut s'écrire :
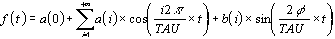
La résolution spectrale décrit les composantes
harmoniques en supposant que chaque variation d'une durée donnée peut être
décrite par une onde d'égale période (harmonique fondamentale) et par des
sous-multiples pairs et impaires de celle-ci (harmoniques secondaires). Les harmoniques constituent
les " formants " tels que le signal peut être reconstruit analytiquement par la
somme des ondes sinus (Fig. 7)
Donc, une résolution en fréquence permet
d'adentifier un signal par sa structure intrinsèque, comme il est probablement impossible que
les propriétés oscillatoires des ondes formantes (" formants "), qui sont
caractérisée par les paramètres amplitude et phase, peuvent appartenir à
un échantillon de variables discrètes ou continues.
En conséquence, l'utilisation de l'analyse spectrale est
la reconnaissance de ce type.
B. Chronobiométrie déductive avec période connue
La Chronobiologie, dans son incessante développement
méthodologique, a été orientée principalement vers analyse de
régression périodique dans laquelle la période d'oscillation est connue
"à priori". Les raisons de cette préférence méthodologique
dépendent du fait qu'en Biologie et en Médecine il est nécessaire de savoir oui
ou non si les fonctions biophysiques ou biochimiques maintiennent la période physiologique dans
le spectre connu des rythmes biologiques (voir Table 1)). Manifestement on doit connaître
"à priori" la période à confirmer. En conséquence, les
méthodes en chronobiométrie déductive avec un période
prédéfinie propose la validation en probabilité de l'hypothèse nulle de
l'amplitude nulle pour une période donnée d'oscillation. Ainsi, toutes les
méthodes montrent l'avantage d'éstimer la signification statistique d'un rythme
recherché (test de sinusoïdalité) qui n'est pas détectable
méthodologiquement en utilisant les méthodes citées ci-dessus.
Plus précisément, le test de sinusoïdalité
est basé sur la probabilité statistique à un niveau P (niveau de détection
d'un rythme) selon que l'affirmation d'amplitude zéro est rejetée. Un rythme est dit
"significatif" ou "non significatif" si P est respectivement < 0.05 ou > 0.05.
L'hypothèse d'amplitude zéro est testée en calculant le Pourcentage Rythmique
"Percent Rhythm (PR)" qui est le pourcentage de variabilité responsable de la fonction
harmonique (rythmique) avec pour référence la variabilité des données
expérimentales égale à 100%.
Les méthodes d'analyse déductive avec
une période connue sont à l'origine de la Chronobiomètrie ou Rythmométrie
descriptive, intégrative et évolutive.
1. Chronobiométrie descriptive (Rythmométrie descriptive)
La Chronobiométrie déductive descriptive
comprend diverses méthodes.
1.1 Analyse du Cosinor
En Rythmométrie descriptive la méthode
la plus importante de régression périodique avec une période donnée est
appelée "Analyse du Cosinor". L'analyse du Cosinor comprend deux méthodes : Le
Cosinor simple et le Cosinor de population. Ces deux méthodes sont basées sur une
approximation sinusoïdale utilisant une fonction Cosinus. Le Cosinor Simple consiste en l'analyse
d'une série temporelle concernant un seul sujet ou un groupe de sujets. Le Cosinor de population donne
plus de détail en sommant les données rythmométriques obtenues par un Cosinor
simple pour plusieurs individus d'un groupe donné.
L'analyse du Cosinor applique comme un opérande
trigonométrique le Cosinus par ce que zéro degré correspond au temps de
référence ou au temps zéro d'un rythme. Le temps zéro coïncide avec
minuit pour les rythmes Circadiens, le solstice d'hiver (le 22 décembre dans
l'hémisphère Nord, 21 juin dans l'hémisphère Sud) pour des rythmes
circannuels.
L'analyse du Cosinor consiste en l'approximation de
l'équation suivante à partir de données expérimentales en utilisant la
méthode des moindres carrés (minimisation) où, M est le Mésor
(acronyme de Midline Estimating Statistic Of Rhythm) c'est le niveau moyen d'oscillation, A est
l'amplitude, mesure de l'oscillation depuis le Mésor ou la moitié de l'oscillation
totale, Pi est 3.14159265, TAU est la période choisie, t est une fraction temporelle du cycle,
une partie de la révolution totale, et Phi est l'acrophase, le temps du maximum de l'onde
(W).
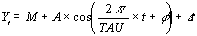
Ainsi, l'analyse du Cosinor permet de mesurer quatre
paramètres de la meilleure onde sinusoïdale calculée : M, A, Phi, W, qui sont
les propriétés d'un rythme pour une période ultradienne, circadienne ou
infradienne.
L'analyse du Cosinor mesure de plus d'autres
paramètres comme le "Percent Rhythm (PR)" et le "Rythm Detection Level (P)". Le profil de
l'onde peut ainsi être validé ou non si l'hypothèse nulle d'amplitude 0 est
rejetée d'une manière significative à un niveau de probabilité
donné. Cela veut dire que le rythme est validé d'une manière significative pour
une période donnée d'oscillation testée par le Cosinor.
La représentation graphique dans un système
d'axes cartésiens de l'onde optimale par la méthode du Cosinor est appelée
"Cosinorgramme" (Fig. 10).
Le traitement du Cosinor permet aussi de représenter
l'amplitude et la phase en coordonnées polaires sur un cercle ou 360 degrés
(sexagésimales) correspondent à la période d'oscillation d'un
phénomène biopériodique. Cette représentation s'appelle un Polargramme
(Fig. 11).
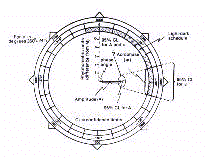
Le Polarogramme permet une description immédiate
des paramètres du Cosinor. L'amplitude et l'acrophase sont décrits en même temps,
et sont représentés par un vecteur partant du centre. La longueur du vecteur est
proportionnelle à l'amplitude, et sa direction indique sur l'échelle temporelle
extérieure l'acrophase en degrés négatifs sexagésimaux ou en temps
physique. Le Polargramme permet la représentation de deux variables à leurs limites de
confiance, la paire amplitude et Acrophase par l'intermédiaire d'une ellipse de confiance
centrée à l'extrémité du vecteur. L'ellipse de confiance montre si un
rythme est significatif ou non, respectivement s'il est situé à l'extérieur ou
à l'antérieur du centre du Polargramme. A noter que les limites statistiques, pour le
Cosinor simple sont données par l'erreur standard de la moyenne (standard error of the mean),
et pour le Cosinor moyenné par les limites de confiance à 95%.
Importance de l'analyse du Cosinor :
L'analyse du Cosinor a plusieurs conditions préalables
fondamentales à l'égard de l'analyse macroscopique des séries temporelles
(temporal series).
La résolution sous forme d'onde élimine les
données fausses ou aberrantes appartenant au bruit biologique ("biological noise").
La fonction sinusoïdale fournie des données rythmométriques qui sont relativement
indépendantes de la densité et de l'équidistance entre les données d'un
échantillon qui est critique en analyse numérique de série temporelle. La
méthode du Cosinor a montré que n'importe quel type d'onde peut être défini
par trois paramètres : mésor, amplitude et acrophase, qui simplifie la description
des rythmes biologiques, et, en plus, facilite la comparaison statistique entre les
biophénomènes oscillatoires. Par contre, l'énorme variabilité du
Chronogramme fait que les séries de données biologiques sont difficiles à
comparer et à décrire dans leur forme périodique. On peut ajouter que les
paramètres rythmométriques fournissent des informations sur les attributs fonctionnels
des phénomènes oscillatoires biologiques. Le profil d'onde, s'il est validé
significatif, démontre que la variabilité temporelle d'un phénomène
biologique donné est systématique et prédictible en parlant de sa période.
Le Mésor fournit une mesure objective du niveau de tonus qui soutient le rythme. L'amplitude,
quantifie le composant phasique qui caractérise n'importe quelle fonction oscillatoire des
organismes vivants. Finalement, l'acrophase estime l'activité périodique dans sa
meilleure expression.
1.2 Analyse spectrale régressive avec périodes connues
Ce type d'analyse est basé sur le principe de
régression périodique avec des périodes pré-fixées qui sont des
sous multiples uniques de celle de l'harmonique principale.
Cela implique que cette analyse est linéaire
en période mais pas en fréquence, exactement le contraire de l'analyse de Fourier.
La formule est (elle manque dans le texte original)
Ou N est le nombre de périodes approchées et
i l'incrément unitaire à partir de la période la plus petite jusqu'à la
période d'harmonique fondamentale. Par exemple, une série de données temporelles
qui couvre 24 heures peut être étudiée dans ses composants harmoniques en validant
le fait que la régression périodique rejette l'hypothèse nulle d'amplitude
zéro à un niveau significatif de probabilité (P < 0.05) pour chaque
période calculée en partant d'un cycle par heure à un cycle tous les 24 heures.
De cette manière, on examine les amplitudes des composants harmoniques unitaires d'un domaine
ultradien ou circadien et leur significativité statistique.
On exprime l'étendue de l'oscillation par le
"percent rhythm". Le spectre est donné par un diagramme cartésien de chaque amplitude
en ordonnée, contre chaque période calculée, en abscisse, ce diagramme est
appelé spectre de puissance "Power Spectrum" (Fig. 12)
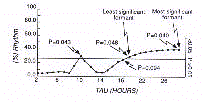
En examinant le spectre de puissance on peut immédiatement
remarquer les harmoniques significatives (" Formants "), comprenant la plus significative
(MSH) et la moins significative (LSH)
En utilisant les harmoniques ("Formants") on peut construire
un modèle harmonique (" harmonic model ") du signal (Fig. 13)
Le modèle harmonique permet la reconstitution du signal
original, non perturbé par le bruit biologique, par l'intermédiaire de la
détermination de l'onde résultante (Fig. 14)
1.2 Analyse en partie de séries ("Serial Section Analysis")
C'est une autre méthode de Chronobiométrie
déductive descriptive dérivée de l'analyse du Cosinor. Cette procédure
consiste en l'itération d'une fonction cosinus sur une série longitudinale de
données biologiques qui couvre un intervalle de temps plutôt qu'une période
d'oscillation (révolution).
Fondamentalement, la méthode applique le Cosinor Simple
(Single Cosinor) à une partie de données séquentielles ("serial section") qui est
incluse dans la période pré-fixée d'analyse (fenêtre chronobiologique).
L'analyse de régression périodique est répétée en décalant
à travers un nombre donné de points de données temporels à
l'antérieur de la fenêtre chronobiologique. La partie sérielle (serial section)
est, donc, composé d'un nombre constant de données dans chaque itération.
L'approche analytique est progressive, et finie quand la fenêtre chronobiologique inclue la
dernière partie des données dans une section sérielle complète
(complete serial section).
Chaque itération calcule les paramètres
rythmométriques, Mésor, amplitude,
Acrophase, et en plus le pourcentage rythmique (percent rhythm) et le niveau de détection
rythmique (rhythm detection level). Par conséquent, l'analyse en section sérielle
(serial section analysis) permet la validation d'une continuité significative des
propriétés rythmométriques sur le temps d'un échantillon. (Fig. 15)
En détaillant, l'analyse évalue oui ou
non 1. Le Mésor et/ou l'amplitude qui à tendance à changer; 2. L'acrophase passe
par un décalage anticipé ou postanticipé ("posticipatory") comme dans la
présence d'un mouvement libre ou des conditions physiques désynchronisées.
1.4 Analyse de régression périodique d'un modèle à plusieurs
composants
L'analyse du Cosinor peut être considérée
comme un modèle de régression périodique à un composant lorsque qu'il
calcule une onde sinusoïdale à travers une série de données temporelles.
Quelques signaux discrets en bio-médecine sont
asymétriques dans leur cycle dans le sens qu'ils montrent une partie différente du temps
dans laquelle les valeurs mesurées sont situées en dessus et en dessous de leur valeur
moyenne. Dans ce cas la meilleure onde calculée est trouvée par les modèles de
régression périodique qui utilisent plus d'un composant harmonique pour approximer des
séries de données temporelles discrètes. Le modèle à plusieurs
composants est habituellement donné par l'onde de période TAU et N sous-harmoniques,
chacune a une période TAU qui est égale à TAU/N.
L'analyse de régression périodique à
plusieurs composants peut s'écrire :
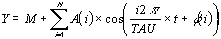
2. Chronobiométrie intégrative (Rythmométrie intégrative)
La Rythmométrie intégrative consiste en la méthode appelée
"Analyse du Cosint"
2.1 Analyse du Cosint
Cette méthode est utilisée pour estimer
les phénomènes bio-périodiques grâce à des fonctions
intégrales par l'intermédiaire de deux limites temporelles de leur période de
révolution.
La méthode du Cosint consiste dans l'analyse d'untégration périodique qui
estime la surface couverte par le profil d'onde le mieux calculé d'un rythme biologique
donné. Les limites t1 et t2 peuvent être égales à la période
étudiée (dans le sens t2 - t1 = TAU) ou à une partie (t2 - t1 < TAU).
Important, l'analyse du Cosint peur être effectuée en connaissant la période
obtenue par régression qui approxime d'une manière optimale la série de
données temporelles. Par conséquent, l'analyse du Cosint peut être appliquer
à des modèles en Cosinus à un composant ou à plusieurs composants.
Dans l'exemple suivant, l'analyse du Cosint est exécutée pour une courbe sinusoïdale
trouvée par l'analyse du Cosinor (Fig. 16)
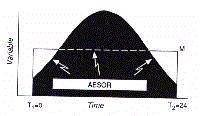
Dans un autre exemple, l'analyse du Cosint est
appliquée à un profil trouvé par un modèle à deux composants issu
d'une régression périodique (Fig. 17)

L'intégral borné par le temps de la courbe
oscillatoire fourni le paramètre Aesor (acronyme pour Area Estimating Statistic Of Rhythm)
L'Aesor est déterminé par la formule suivante,
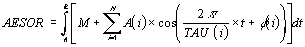
qui est relative à la fois à des
modèles mono-composants (méthode du Cosinor) et multi-composants, d'analyse de
régression périodique (voir au dessus)
La surface de l'intégrale est calculée en
sommant les sous surfaces dans lesquelles
la surface peut être divisée en distances unitaires de base de temps (unité de
temps) correspondant à la période du rythme examiné (par exemple, la minute pour
un rythme Circadien, le jour pour un rythme Circaseptan, le mois pour un rythme Circannuel). La
distance unitaire en temps correspond à un intervalle h. Les sous surfaces sont données
par les surfaces quadratiques dont les cotés correspondent aux deux valeurs de Y
calculées par la fonction de régression périodique dans l'intervalle h comme
base de temps (Fig. 18)
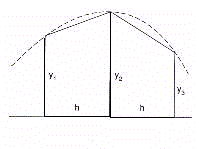
La surface des deux sous-surfaces est donnée par la
formule :
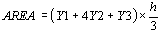
La surface totale est obtenue par la somme de toutes les sous surfaces unitaires (Fig. 19)
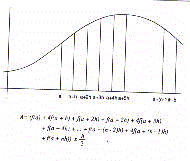
L'estimation de la surface totale, l'Aesor, est ainsi donnée par la formule suivante :
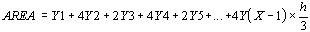
Vu que les valeurs de Y1, Y2, Y3, Y4 sont données
par la fonction de régression périodique F(Y), la formule devient :

3. Chonobiométrie évolutive
L'analyse des cycles avec variations temporelles
peut être séparée en deux méthodes qui respectivement pour la
première analyse les changements du Mésor, de l'amplitude et de l'acrophase, et pour
l'autre de la période.
3.1 Analyse du Clinospectror
Ce type d'analyse est relatif aux changements
du Mésor, de l'amplitude et de l'acrophase (mais pas de la période) de rythmes
biologiques.
Jusque là, les méthodes déductives
qui ont étés décrites apparaissent comme capablent d'axaminer et de
décrire un phénomène biologique à un temps donné d'un cycle de vie.
On doit remarquer, pourtant, de nouveau que la majorité des rythmes biologiques ont tendance
d'avoir leurs propriétés oscillantes qui varient lorsqu'illes vieillissent.
La Rythmométrique appliquée lors de vieillissement affecte le Mésor,
l'amplitude et l'acrophase (Gerorythmométrie). Il y donc un besoin d'étude
biométrique du fait des changements complexes en regard de la connaissance des
procédures appropriées (Clinorythmométrie). L'approche des changements relatifs
à l'âge dans les paramètres rythmométriques nécessite une analyse
de régression périodique linéaire dans laquelle la variation des
paramètres périodiques est analysée par une fonction linéaire de
l'âge. Ainsi on utilise une méthode appelée analyse du Clinospector. La formule
est la suivante :
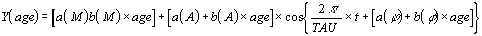
Où a est le retard ("intercept"), c'est à dire,
la valeur des paramètres du rythme à l'âge 0, et b est le coefficient angulaire de
la direction (trend), et M, a et Phi sont les paramètres Rhytmométriques, t est une
fraction du temps de la période étudiée.
La régression périodique linéaire
optimale est déterminée par l'analyse du Clinospector représentée par le
Clinorythmogramme, un graphique plan ou l'ordonnée et l'abscisse sont respectivement
l'oscillation autour du Mésor et l'âge chronologique. Des exemples du Clinoryhtmogramme
sont donnés par la figure 2.
Selon la Clinorythmométrie, plusieurs modèles
de vieillissement peuvent être idendifiés comme nous le verrons dans les chapitres
suivant.
En outre, l'analyse du Clinospector peut être utilisée pour prévoir
l'évolution d'un rythme biologique comme une fonction du cours de la vie. Il est possible ainsi
de faire des projections dans des temps futurs en plus d'extrapolations dans le passé
jusqu'à la naissance. Il est aussi possible de prévoir un âge biologique
(BA : Biological Age) en tenant compte de l'âge chronologique (CA : Chronological
Age), et de calculer le coefficient de sénilité (SI : Senility Index) à
partir de la formule : SI = BA / CA.
3.2 Méthode de démodulation complexe
Les rythmes biologiques peuvent avoir une période
qui varie en fonction de l'âge chronologique. L'âge relatif change en fréquence et
peut être estimé par le modèle de démodulation complexe suivant
l'équation :
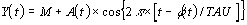
Il est important d'insister sur le fait que la méthodologie en Chronobiologie est
composée par d'autres procédures relatives à d'autres analyses. En plus, la
Chronobiologie utilise d'autres standards de référence, comme, le Chronodesms, le
Cosinordesms, l'Aesoredesms. Le but de cet article est de voir la méthodologie en
Chronobiologie dans ses différents aspects. Les lecteurs peuvent avoir un "stimulus" en consultant
les publications citées plus loin, s'ils veulent plus de
détails.
CONCLUSIONS
Cet article est une tentative concise pour tracer
les grandes lignes des principes et des
méthodes en Chronobiologie. Pour plus d'information les articles qui composent cette
publication sont fournis. Ces publications peuvent être considérées comme un
"summa chronobiologica" (le summum en Chronobiologie) étant donné qu'ils ont
été écrits par le Chronobiologistes les plus éminents
dans le monde.
BIBLIOGRAPHIE CONSEILLEE
Ajello L. e Fea F. I Bioritmi. Editrice Minerva Medica, 1959.
Arendt J., Minors D.S. and Waterhouse J.M. Biological Rhythms in Clinical Practice. Wright, London, 1989.
Aschoff J. Zeitgeber der tiorischen Tagesperiodik. Naturwissenschaften 41: 1954: 49-56.
Aschoff J. und Wever R. Naturwissenschaften 49: 1962: 337-342.
Aschoff J., Daan S. and Groos G. Vertebrate Circadian Systems: Structures and Physiology. Springer-Verlag, Berlin-Heidelberg-New York, 1982.
Bingham C., Arbogast B., Cornelissen G., Lee J.K. and Halberg F. Inferential statistical methods for estimating and comparing cosinor parameters. Chronobiologia 9: 1984: 397-442.
Biological Rhythms in Clinical and Laboratory Medicine. Y. Touitou and E. Haus (Eds). Springer-Verlag, Berlin-Heidelberg- New York, 1992.
Carandente F. Elementi di Cronobiologia Sperimentale e Clinica. Il Ponte, Milano, 1990.
Chronobiology: Basic and Applied. In: P. Cugini, F. Halberg, D.W. Wilson and P. Pasquini (Eds). Ann. Istit. Sup. San. Vol. 29. Istituto Poligrafico e Zecca dello Stato, Roma, 1993, pp. 481-710.
Colquhoun W.P. Aspects of Human Efficiency: Diurnal Rhythms and Loss of Sleep. English Univ. Press, London, 1972.
Cugini P. Perché la Cronobiologia. La Nuova Critica, 1981, pp.93-119.
Cugini P. Dynamic rhythmometry: Inferential excess indices using microcomputers. J. Interdisc. Cycle Res. 18: 1987: 297-306.
Cugini P. Istituzioni di Cronobiologia Medica. Introzzi, USES Edizioni Scientifiche, 1988, pp. 1729-1806.
Cugini P., Lucia P., Murano G., Di Palma L., Battisti P. e Cogliati A. Cronobiologia per la medicina: principi e metodologia. Agg. Med. 12: 1988: 675-685.
Cugini P. Chronobiologic Approach to Aging: the Clinospectroscopic Method. Style Compostampa srl, Roma, 1991.
Cugini P, Di Palma L. Cosint analysis: a procedure for estimating biological rhythms as integral function by measuring the area under their best-fitting waveform profile. Biol. Rhythm Res. 25: 1994: 15-36.
Endocrine Rhythms. D.T. Krieger (Ed). Raven Press, New York, 1979.
Folkard S. and Monk T.H. Hours of Work. John Wiley and Sons, Chichester, 1985.
Gedda L. e Brenci G. Cronogenetica: L'eredità del Tempo Biologico. Edizioni Scientifiche e Tecniche, Mondadori, 1974.
Halberg F. Tong Y.L. and Johnson E.A. Circadian system phase: an aspect of temporal morphology-procedures and illustrative examples. In: The Cellular Aspects of Biorhythms. Von Mayersbach H. (Ed). Springer, Berlin 1967, pp. 20-48.
Halberg F., Carandente F, Cornelissen G. and Katinas G.S. Glossary of Chronobiology. Chronobiologia, 6 (suppl.1): 1977.
Halberg F. I ritmi biologici nella pratica clinica. Minuti 5: 1978: 27-37.
Halberg F., Lee J.K. and Nelson W.L. Time-qualified reference intervals, chronodesms. Experientia (Basel) 24: 1978: 713-716.
Harris R.G. Biological Clocks. Cold Spring Harbor Symp. Quant. 1986, pp. 85-113.
Hastinggs J.V. and Schweiger H.G. The Molecular Basis of Circadian Rhythms. Dahlem Konferenzen, Berlin, 1976.
Hekkens W.Th.J.M., Kerkhof G.A. and Rietveld W.J. Trends in Chronobiology. Pergamon Press, Oxford, 1988.
Hickey D.S., Kirkland J.L., Lucas S.B. and Lye M. Analysis of circadian rhythms by fitting a least squares sine curve. Comp.Biol. Med. 14: 1984: 217-223.
Hildebrandt G. Biologische Rhythmen und Arbeit. Springer-Verlag, Wien-New York, 1976.
Manfredini R., Gallerani M. e Fersini C. Cronobiologia dell'apparato cardiovascolare. Editoriale Fernando Folini, Casalnoceto (Alessandria), 1995.
Minors D.S. and Waterhouse J.M. Circadian Rhythms and the Human. Wright-PSG, Bristol, 1981.
Nelson W., Tong Y.L., Lee K and Halberg F. Methods for Cosinor-rhythmometry. Chronobiologia 6: 1979: 305-323.
Nelson W., Cornelissen G., Hinkley D., Bingham C. and Halberg F. Construction of rhythm-specified reference intervals and regions, with emphasis on 'hybrid' data, illustrated for plasma cortisol. Chronobiologia 10: 1983: 179-193.
Nicholson A.N. Sleep, Wakefullness and Circadian Rhythm. AGARD Lecture Series No. 105, 1979.
Orologi Biologici. Le Science. (Quaderni) 9: 1983: 2-79.
Palmer J.D., Brown F.A.Jr. and Edmunds L.N. Introduction of Biological Rhythms. Academic Press, New York, 1976.
Redfern P.H., Campbell I.C., Davies J.A. and Martin K.F. Circadian Rhythms in the Central Nervous System. MacMillan Press LTD., London, 1985.
Reinberg A. Dai Ritmi Biologici alla Cronobiologia. I Quaderni de 'Il Ponte'. Chronobiologia (Suppl.2): 1975: 7-97.
Rietveld W.J. Clinical Aspects of Chronobiology. Meducation Service Hoechst, 1986.
Sanchez De La Peña S., Halberg F., Halberg E., Ungar F., Cornelissen G., Sanchez E., Brown G., Scheving L.E., Yunis E.G. and Vecsei P. Pineal Modulation of ACTH 1-17 Effect upon Murine Corticosterone Production. Brain Research Bulletin 11: 1983: 117-125.
Sensi S. Elementi di Cronobiologia Clinica. Esculapio Ed., Bologna, 1984.
Tarquini B. Cronobiologia Pratica. Esculapio Ed., Bologna, 1984.
Timing and Time Perception. J. Gibbon and L. Allan (Eds). Ann. N.Y. Acad. Sci. 423: 1984.